引 言
2020年4月8日《细胞》杂志、2020年6月24日《自然》杂志分别在线发表了上海神科所研究员杨辉团队和加州大学圣地亚哥分校教授付向东的论文:两篇论文用不同的手法敲低同一个分子(PTBP1),成功在小鼠脑内将胶质细胞转化为有功能的神经元—— 这对 “再生疗法” 用于治疗神经退行性疾病是一个重要的里程碑进展。2020年7月2日,付向东实名举报杨辉涉嫌剽窃其工作以及造假等学术道德不端行为的举报信在知乎等网络平台得到广泛传播,其中提到他为鉴定证明PTBP1的作用投入了9年多的时间和工作,包括 6 年的实验工作,以及近 3 年审稿过程中的补充实验工作。
那么,这一过程是什么样的呢?本文基于6月24日对付向东及其实验室博后钱浩博士的采访,对他们围绕PTBP1所做的科学探索之路进行了梳理。

2009年年底。
看着显微镜下海拉细胞表面长出的枝丫,正在为博士后出国训练做准备的薛愿超感到十分不解。
海拉细胞是一类表皮细胞,在1951年由约翰霍普金斯医学院的研究者从一位名叫海莉·拉克斯的子宫颈癌患者中分离培养出的永生细胞系。在半个多世纪的时间里,海拉细胞在条件适宜的环境下平均每22小时数量翻倍一次,产生的新细胞的形态也与上一代高度相同:凭借着像梭子一样扁平化的细胞结构,它们能紧贴在培养皿中底部旺盛生长,那由磷脂双分子层构成的细胞边缘十分平整,给人一种流线型设计的美感。
全世界各个实验室储存的海拉细胞数以亿计,大都保持着和半个多世纪以前那批子宫颈癌细胞几乎一致的形态。
但展现在薛愿超眼前的这盘海拉细胞,却在原本平滑的表面上多出了许多的分叉。这就好像是在斑马头上长出鹿角之类的配件一样诡异。他清晰地记得当时的情况,大约两周前他把这盘细胞放进恒温培养箱,细胞的表面绝对不是这样的。这个培养皿中的细胞原本应长得和其他亿万个海拉细胞一样,梭子状轮廓的边缘十分光滑。
薛愿超不禁开始思考:在过去两周的时间里,这些细胞内部究竟发生了怎样的变化?
这些海拉细胞系的特别之处,是薛愿超在它的基因组中插入了一段转基因序列,使得其可以表达一小段 RNA 序列。这个精心设计的 RNA 就像是一颗 “定时炸弹”,可以基于碱基互补配对选择性地与细胞中一个名为 PTB 基因的 RNA 结合—— 一旦结合,便会引爆,吸引细胞内的 RNA 降解蛋白将 PTB 基因的 RNA 清除,没了 PTB 基因的 RNA,细胞自然也无法再大量合成出 PTB 蛋白。
生物学家研究一个问题,通常会从充分性和必要性两个角度进行研究。充分性往往是在一个系统中人为增加一些东西,看看会引起哪些变化;必要性则是去人为地减去系统中的某些成分,其中最常用的手段就是通过基因编辑或者 RNA干扰的 “定时炸弹”,精确地下调某一个基因产物的水平。
薛愿超为什么想要下调海拉细胞中的 PTB 蛋白量呢?我们后面会提到。
听了学生汇报海拉细胞长出奇怪枝丫的观察,加州大学圣地亚哥分校教授付向东内心忽地划过一道闪电,意识到这背后很可能隐藏着一个重要的科学发现。斑马的头上长角很奇怪,但是如果这对角长在鹿的头上,就不会让人感到奇怪。这种枝丫的结构,在表皮细胞中稀奇,但是神经细胞中却非常普遍。在海拉细胞中敲低 PTB 蛋白,会不会像是对斑马施展了一道神奇魔法,让它头上长出鹿角了呢?
这是付向东的直觉思维,接下来他想到了近期分子神经生物学领域的几个最新研究:
第一,PTB 蛋白在动物细胞中还有两个高度相似的 “兄弟姐妹”,其中之一就是 nPTB:它们的 DNA 序列高度保守,不同之处在于 PTB 在几乎所有的细胞中都大量表达,而 nPTB 仅仅局限在神经细胞中表达(nPTB名字中的n的意思就是neuronal,即神经的);[1,2]
第二,nPTB 的蛋白与许多参与神经发育的因子有关联,nPTB 可能是一个触发细胞向神经元分化的 “开关”;
第三,PTB 会负向调节 nPTB 蛋白产物的表达量:至少在当时已经发现PTB的蛋白产物可以结合 nPTB 的 RNA,通过促进 nPTB 的 RNA 的降解,下调细胞中 nPTB 的蛋白量。
基于此,付向东觉得这个奇怪的发现很有意思,他建议薛愿超追着这条线索往下继续侦察下去。
于是,一段奇妙的科学探险就此拉开帷幕。而此前付向东作为一名分子生物学家,研究 RNA 研究将近四十年,这是他了解 RNA 剪切、PTB 以及神经再生疗法的关键。
自从1983年付向东通过中美生物化学联合招生项目(CUSBEA)来到美国凯斯西储大学,师从 Jonathan Leis 研究鸟类逆转录病毒的增殖调节开始,RNA 就成了他科学生涯的核心。
RNA 全称核糖核酸,在维持细胞正常功能中扮演重要角色:它们在细胞核内以 DNA 为模板转录产生,然后在细胞质中发挥多种多样的作用。正因为其功能对于细胞的生存异常重要,细胞演化出了一系列能够调节 RNA 序列组成和稳定性的精密机制。但如果某个基因的 RNA 数目超出了细胞的需求,细胞还会启动一套特殊的 RNA 降解分子机器,通过主动清除冗余的 RNA,保证细胞内 RNA 水平刚好能够满足细胞需求。这套被称作 “RNA干扰” 的分子机制的发现,2006年得到了诺贝尔生理学或医学奖的认可。
在这一领域,付向东于1990年代鉴定出的 RNA 剪切因子 SC35、第一个 RNA 剪切因子激酶,推进了人们从分子生物学层面对RNA剪切过程的认识。
然而,21世纪初的国内学术界,RNA 生物学的研究水平与国际顶尖力量有不小的差距。因此,接到昔日同窗、武大生科院负责人发出的回母校——武汉大学任教的邀请时,付向东觉得这是一个领导推动国内 RNA 生物学领域前进的良机。
付向东选择了兼职,成为武大的咨询顾问,与武大教授张翼共同指导研究生。回到武汉大学,他不但能有机会多看望居住国内的父母,还能带动国内一个领域的发展,这无疑是促使付向东作出回国决定的两个重要考虑因素。此外,受益于 CUSBEA 项目培养,他亦将推动国内生物学研究生教育视为自己的一份责任。
在2004年入学的博士研究生招生过程中,一位姓薛的年轻人引起了付向东的注意。在众多向往武汉大学生命科学学院博士研究生的申请者中,本科毕业于信阳师范学院的薛愿超的简历并不耀眼,但付向东并没有将本科学习成绩当作招收研究生的唯一评判标准:他给申请者们发了一篇近期发表的Science论文,要求大家写下读后的感想。
在交上来的作业中,有一份特别的阅读报告,对问题的思考很有深度,一下子吸引了付向东的注意。薛愿超如愿被武汉大学录取,在张翼和付向东的联合指导下攻读博士学位。
在薛愿超加入付向东和张翼的联合团队时,RNA生物学领域的科学家们已对 RNA 结合蛋白产生了浓厚的兴趣:在当时,已经有不少证据提示 RNA 结合蛋白的功能和神经性疾病、自身免疫病存在密切联系。但在分子机制水平,这些 RNA 结合蛋白究竟靶向哪些特殊的 RNA 分子仍然存有巨大的谜团。付向东和张翼也跃跃欲试,带领团队加入搜索 RNA 结合蛋白靶点的寻宝大军中。
相比于 DNA,RNA 有更丰富的二级结构,且更不稳定,这增加了研究 RNA 结合蛋白靶点的难度。2003年,洛克菲勒大学 Robert Darnell 在方法学上发明的 CLIP 技术,为人们寻找 RNA 结合蛋白所结合的 RNA 分子铺平了道路。CLIP 新技术对于想要深入研究 RNA 结合蛋白的付向东而言,如同一柄削铁如泥的宝剑。
付向东先按照 Robert Darnell《科学》论文中公布的配方,在自己的实验室照葫芦画瓢,锻造出一把锋利的仿制品,并将这项 “打铁” 的任务交给了动手能力极强的薛愿超来做。宝剑出鞘,该使出怎样的剑招呢?他们需要确定运用 CLIP 技术研究的对象。
在细胞内数量众多的 RNA 结合蛋白当中,他们将目光聚焦到了一个名为 PTB 的蛋白身上。
PTB全称多聚嘧啶区结合蛋白(polypyrimidine tract-binding protein),得名的原因来自最初克隆它的 Philip Sharp 教授团队观察到其能结合富含嘧啶的碱基序列。Sharp 团队最初推测其是一个能直接介导 RNA 剪切因子(splicing factor),但后来其他团队的工作表明 PTB 其实是一个参与调节 RNA 剪切反应抑制性因子(splicing repressor)。付向东很擅长 RNA 剪切领域的研究,再加上实验室中正好有高度特异性的 PTB 抗体可用,他们就决定将 CLIP 技术首先应用于 PTB 靶点 RNA 的搜寻。[4-6]
薛愿超博士五年工作的关键成果,就是在全基因组水平鉴定了HeLa人细胞系内可能与 PTB 蛋白结合的 RNA,该发现于2009年发表于《分子细胞》杂志。PTB 蛋白家族包含了三个成员:除了在大部分体细胞中都广泛表达的 PTB 之外,还有局限于神经细胞中表达的 nPTB 和在免疫细胞中选择性表达的 ROD1。因此,在推进 PTB 课题的同时,薛愿超也利用 CLIP 技术在寻找 nPTB 作用的 RNA 序列。
然而,相比于 PTB,nPTB 结合靶点课题就好像是一份在地下埋藏得更深的宝藏,这主要因为不同于普遍表达的 PTB,nPTB 局限于神经细胞中表达,而且仅仅在神经元发育的早期表达。就像是捕鱼,PTB 蛋白在各个池塘中都有,且全天候活动;而 nPTB 只在神经细胞这片 “池塘” 中存在,且在特定的一小段时间内抛头露面,这无疑增加了垂钓者的难度。探索 nPTB 蛋白结合的RNA 靶点,如果仅仅直接照搬《分子细胞》论文中的方法,还行不通。
薛愿超需要对实验条件进行优化。2007年,UCLA 的 Douglas Black 团队发现,PTB 蛋白能够结合在 nPTB RNA的内含子区以调节剪切位点的利用,导致 nPTB 转录本的第十个外显子在 RNA 剪切加工过程中被跳过,使得其在细胞中被快速降解,抑制细胞中 nPTB 蛋白含量。[1]
如果把 PTB 和 nPTB 想象成两条鱼,假设前者会以后者为食进行捕捉,致使后者大部分时间都躲在暗处,只有在一部分 PTB 水平较低的细胞中,nPTB 才会选择在特定的时间段露面。
要想运用 CLIP 技术寻找 nPTB 的结合靶点序列,需要让 nPTB 蛋白在胞内稳定地达到足够高的浓度。于是很自然地读者通常能想到的第一个法子是在细胞系中对 nPTB 进行过量表达,但在此处由于 PTB 是在 RNA 水平抑制 nPTB 的表达,那么即使表达了更多的 nPTB,PTB 也会兵来将挡,细胞中依然没有足够多的 nPTB。
于是,薛愿超想到了第二个办法:他决定使用RNA干扰技术,对细胞内的 PTB 蛋白进行下调。他的思路是,如果平日 “欺压” nPTB 的 PTB 被他铲除了,nPTB 蛋白就敢冒出头来。一旦 nPTB 蛋白在细胞中稳定存在,那么他就能利用 CLIP 技术在全基因组范围内寻找 nPTB 的 RNA 靶点。
他设计了一个 shRNA 质粒,使得其在插入海拉细胞基因组后能稳定大量表达靶向 PTB 的 “定时炸弹”。这就是开头中出现的那个长出枝丫的奇怪细胞系。
付向东和薛愿超都对这个意外的发现感到十分惊奇。于是在博士毕业后,薛愿超决定到付向东位于圣地亚哥的实验室,在博后阶段继续探索 PTB 敲低的海拉细胞中长出枝丫的神秘原因。
来到圣地亚哥以后,薛愿超首先在多个不同的细胞系中通过 RNA 干扰技术稳定敲低 PTB 表达量—— 多个细胞系均出现了枝丫状的轮廓,肯定了他此前的观察不是一个偶然的特例。新转化形成的细胞的基因表达也发生了变化,许多在神经细胞中特异性表达基因的表达量在 PTB 下调之后也显著增加了。
更让他感到高兴的是,在小鼠胚胎成纤维细胞(MEF)中敲低 PTB,细胞不仅在形态上长得更像神经细胞了,而且进一步电生理实验结果表明,新转化得到的细胞在功能上表现出与神经细胞相似的特性—— 可以产生动作电位(action potential)。这些结果都提示,降低细胞中 PTB 蛋白表达量,能诱导成纤维细胞分化为一类近似于神经细胞的细胞类型,说明在细胞命运决定的过程中,PTB 可能是一个阻断细胞向神经细胞转化的“路障”。
但作为一个调控 RNA 剪切过程的蛋白,PTB 是如何影响细胞命运的决定呢?付向东和薛愿超需要在机制上为自己的发现找到一个合理的解释。
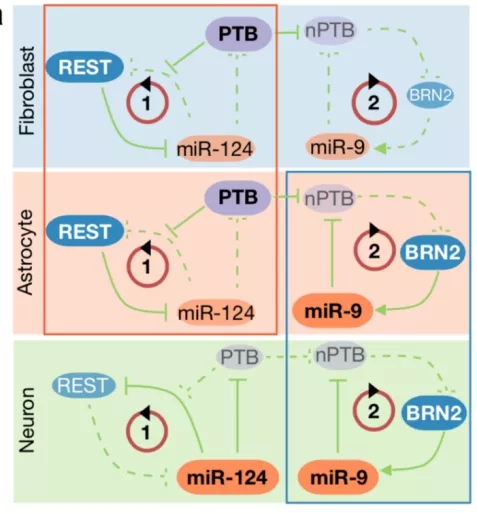
他们推测,PTB 诱导细胞向类似于神经细胞方向分化,可能是通过 RNA 剪切调节了与神经细胞分化相关的转录因子。在当时已经报道的一打的诱导神经细胞发育的转录因子当中,一个名为 REST 的蛋白吸引了他们的注意力:
第一,REST 蛋白顾名思义,是让细胞维持在一种 “休息” 的状态,是一道阻止细胞向神经元分化的屏障。
第二,REST蛋白已经被发现存在不同的同源异构体(splicing isoform),即REST基因的成熟mRNA中存在多种外显子的组合方式。这意味着REST的表达量很可能在RNA剪切过程中经历了关键的调节。
第三,已知REST蛋白在被miR-124特异性结合后会快速降解。
经过严谨的细胞生物学实验,付向东和薛愿超最终证明 PTB 也能结合 REST及其复合物中多个关键组份,从而阻止其经历 miR-124 介导的降解,使得细胞中 REST 转录因子维持在较高的水平,于是细胞难以向神经细胞方向转化。这项发现最终被发表于2013年的《细胞》杂志,引起了业内广泛的关注。[8]
这是因为如果真的通过调节一个 RNA 结合蛋白就能诱导产生新的神经元,那么在众多因为神经元死亡而导致的神经退行性疾病中,都有可能通过阻断 PTB 通路诱导产生新的神经元作为治疗的手段。PTB 在神经细胞命运决定中的作用为神经退行性疾病的 “再生疗法” 带来了新的曙光。
付向东的团队以分子生物学手段见长,但对于神经生物学实验的经验并不多。因此,尽管早在2011年他们就已经开始尝试在体内通过下调 PTB 诱导产生新的神经元,但进展却比较慢。
PTB介导的神经元转分化通路模型[9]
就在同一年,中科院生物物理所神经生物学家王晋辉培养出的一位优秀博士研究生钱浩加入了付向东的团队,挑起了实验室神经生物学实验的大梁。那么,基于 PTB 的再生疗法能成为治疗神经退行性疾病的 “天降神兵”,还是说一切到头来只是竹篮打水一场空?
2013年,RNA 结合蛋白 PTB 控制神经细胞命运决定的发现,燃起了人们对于神经退行性疾病再生疗法的热情和期待。但从基础研究走向临床的道路有太多的阻碍,这篇论文的发表仅仅是万里长征的第一步。
付向东和薛愿超对于这一点心知肚明。首先摆在他们眼前的问题,是搞清楚人和小鼠细胞命运决定通路之间的差异。他们发现,尽管小鼠成纤维细胞在敲低 PTB 后可以分化成在形态上与功能上都类似于神经细胞的类型,但人的成纤维细胞在同样敲低 PTB 之后却仅仅在形态上 “形似”,却不具有神经细胞所具备的诸多电生理学性质。
在体外得到的神经细胞仅仅 “形似” 而 “神不似”,空长了一副接近神经细胞的皮囊,却没有神经细胞的功能—— 如果在这种情况下贸然直接进行体内实验,纵使诱导出了大量形似神经的细胞,由于缺乏功能,对于疾病的帮助可能也极为有限。
因此,他们的当务之急是找出为什么敲低 PTB 在小鼠中可以得到具有功能的神经细胞,而在人的成纤维细胞中却只能产生出一堆无功能的细胞。这个问题就像是悬在付向东和薛愿超头顶的达摩克利斯之剑。
在人成年成纤维细胞中,敲低 PTB 能引起 nPTB 产生,细胞呈现出类似于神经细胞的外观,表明很可能已经诱导向神经细胞方向分化,但在中间停了下来。如果将人成纤维细胞向神经细胞分化的过程想成是一块巨石从山坡滚到山脚的过程,那么敲低 PTB 在外观形态学上引起的改变可能已经 “推动” 了巨石向下的运动,但在到达半山腰的某处却因为有障碍物而被拦了下来。这个 “路障” 在小鼠中阻力较小,因此决定细胞命运的巨石凭借自身强大的惯性得以平稳通过;但在人成纤维细胞中内在的阻力较大。
付向东和薛愿超设想,如果能找到挡道的 “路障”,将其清除以后,人细胞命运的巨石就也能有始有终,最终分化为形态与功能兼具的神经细胞。他们将从山顶到半山腰的细胞命运决定过程称作 “神经诱导”(neural induction),将半山腰拦路石与山脚之间的部分称作 “神经成熟”(neuronal maturation)。这就是 PTB 介导的神经细胞分化的 “两步走模型”(2-step model)。
那么,人成纤维细胞中的半山腰处的这块 “拦路石” 究竟是什么?为了研究这块 “拦路石”,付向东和薛愿超采取了一个巧妙的思路:已知转录因子在神经细胞分化中扮演着类似于 “钥匙” 的重要作用,并且研究者也已经报道了一系列这样的转录因子。那么 PTB 下调也应该是通过一些神经细胞发育相关的转录因子帮助细胞顺利通过半山腰的拦路石。只要找出移除这些障碍的转录因子,就距离目标更近了一步。
2016年,他们终于筛选到了一个关键的转录因子—— BRN2。这个发现于2016年发表在《自然神经科学》。[10] 但更让付向东团队兴奋的,是来自于钱浩在星形胶质细胞体外培养实验中所作出的观察。
星形细胞是在成熟大脑内大量存在的一类胶质细胞。不同于已经停止分裂的神经细胞,星形细胞在接收到生长信号后依然能快速复制增殖。因此,试图将星形细胞转变为神经细胞,是神经退行性疾病再生疗法中最具有潜力的策略之一。
就在薛愿超开展寻找 BRN2 实验的同时,钱浩在薛愿超和胡婧提供的分子生物学技术支持下,通过 RNA 干扰技术敲低体外培养的星形细胞中的 PTB 表达量,发现体外培养下的星形细胞可以转化为在形态、基因表达和电生理性质都接近于神经细胞的细胞类型。
原来,在星形细胞中,BRN2 始终处于较活跃的状态。换句话说,由于 BRN2 一直存在,星形细胞向神经细胞分化的半山腰上的 “拦路石” 拦不住分化的势头了。这真是实在是天公作美,钱浩立刻开展体内了实验,探究 PTB 敲低在生理学环境下的影响。
但付向东和钱浩在开展体内实验时,对于实验结果并不乐观。此前也有一些实验室报道可以通过控制特定转录因子的表达能够在体外诱导产生神经元,但是这些靶点一旦到了环境更为复杂的体内,却纷纷失灵。[11-13]
生物医学的研究有的时候真的就像是魔法,虽然中间还有许多未解的黑箱,其中任何一个环节出错都会让整套实验的希望化为泡影。但也有少数的实验,听上去困难重重,但竟然真的能够像走钢丝一样不出差错地收获成功。
钱浩的体内实验很快取得了进展。他向中脑区域注射能敲低 PTB 水平的 AAV 病毒,中脑中一部分星形细胞形态变化为神经细胞的样子,表达出神经细胞特有的基因,获得了神经细胞特有的电生理性质,并且伸出长长的轴突直达纹状体。
此外,他还向帕金森模型小鼠的中脑区域注射 AAV 病毒敲低 PTB,小鼠运动能力显著恢复,提示新转化出的神经元具有参与运动控制的功能。
反义寡核苷酸是目前比较接近临床应用的基因治疗手段,已经有治疗神经系统疾病的 ASO 药物在国内外获批上市。为了向临床方向努力,钱浩化学合成了靶向 PTB 的 ASO 药物,注射入帕金森模型小鼠体内,小鼠运动能力得到修复,概念验证性实验结果喜人。
2017年12月,付向东将这些结果整理成文,投到了《科学》杂志。这些结果如果能得到更多团队独立的验证,无疑将会成为神经疾病再生疗法的重要里程碑。付向东抑制不住内心激动的心情,甚至在投稿到《科学》之前,就已经开始在一些学术研讨会和会议报告中与同行分享这奇妙发现的喜悦。这些结果看上去如此完美,究竟是真是假?
科学家偶尔会被自身的狂热冲昏头脑,由于不识庐山真面目,对实验数据进行过度的解读。因此,学术共同体遏止这种 “科研大跃进” 的一个关键措施就是 “同行评议” 制度,如同一台X光照相机一样对论文的逻辑漏洞逐一检查。论文的作者只有对审稿人提出的问题进行合理的回答,杂志的编辑才会最终接受、发表论文。
付向东关于靶向 PTB 蛋白进行神经细胞再生疗法的论文看上去过于完美(too good to be true)。对于这个发现,杂志编辑和审稿人有责任用苛刻的眼光对文中的结论进行检验。这种检验尽管有时候会被认为是 “鸡蛋里挑骨头”,大大放慢了科学论文发表的速度。
在综合了几位审稿人的意见后,《科学》杂志的编辑,觉得尽管这项工作意义重大,但是仍然对实验的可重复性十分担忧,因此拒收这篇论文。尽管遭到了《科学》杂志的拒稿,付向东和钱浩依然得到了很多建设性的建议。经过一些修改,他们于2018年11月将工作投稿到了《自然》杂志。
付向东提交给《自然》的论文草稿吸引了编辑们浓厚的兴趣—— 毫无疑问,如果付向东的这个发现能被其他团队稳定重复,绝对会是一个里程碑式的突破。但越是重量级的成果,越需要小心谨慎。《自然》对付向东的这篇投稿非常重视,邀请了四位审稿人进行同行评议。审稿人无一例外的对这项工作的重要意义给予了充分的肯定,但同时也就其中包含的漏洞提出了许多建设性的意见。
《自然》杂志的编辑在发给付向东的邮件中说,只要他们能合理地就审稿人们提出的问题给予合理的答复,《自然》一定会发表这项工作。他们不用担心被其他团队抢发的风险,因此有充足的时间可以安心地将文章的关键结论做得尽可能扎实。
在审稿人围绕论文结论提出的众多问题中,最关键的问题有三个:
第一,基于付向东团队所展示的数据,他们无法说明在帕金森模型小鼠中,是新转化产生的神经元直接导致行动能力的恢复。因为他们的实验结果只能说明在中脑注射 AAV 病毒敲低 PTB,能观察到行为能力的恢复——对于这一结果还存在另一种可能的解释,即感染鼠脑的 AAV 病毒通过影响其他本来存在的神经元的状态恢复运动能力。
第二,付向东的投稿中对于新转化产生的神经元具有完整功能的证据尚不十分充分。付向东观察到由中脑星形细胞转化形成的神经元可以将突触投射到纹状体核团,但这仅仅是结构角度的证据。审稿人认为要想坐实关于由转化产生新神经元具有功能的结论,需要在对这部分神经元进行刺激后,在纹状体区域记录到真实的神经递质释放。
第三,小鼠和人的大部分神经退行性疾病有更大的概率发生在年老体衰之时,而细胞的分化能力会随年龄的增加而逐步降低。投稿中付向东展示的实验主要为年轻鼠的数据,而神经元再生疗法成立的一个重要基石是在年老人群中也可以诱导产生新的神经细胞。因此,审稿人认为付向东至少应该在年老小鼠中检验转分化策略的治疗效果。
这三个问题都问得一针见血,再加上编辑们作出的不担心被抢发的保证,付向东和钱浩打算静下心来慢慢打磨这篇文章,在把审稿人要求的主要实验都完成后再将论文投回到《自然》。相比于急着发表出来彰显自己的成果,精益求精地把结论做到尽可能的扎实完整会更有意义。
仔细阅读审稿人的反馈意见之后,钱浩和付向东一起讨论应对三个核心问题的对策:
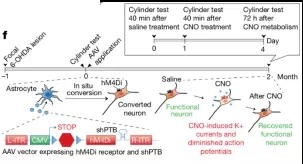
对于问题一,他们设计了一个基于化学遗传学的精巧实验:在装载了表达 shPTB 的 AAV 病毒载体上,他们利用分子生物学同时表达了一个人工改造的G蛋白偶联受体(G-protein coupled receptor(GPCR))—— Gi-DREADD,这个特殊的G蛋白偶联受体一旦表达在细胞膜上,就会在人工注射药物 CNO 时被激活,导致细胞变得更加难以被激活。
于是,他们根据此前的实验条件诱导星形细胞转化为神经细胞,并使得这些转分化的神经细胞表达化学遗传学受体。他们首先观察到帕金森模型小鼠的异常行为因为这些转分化神经元的作用而得到恢复;随后再用配体药物 CNO 激活这些受体,转分化的神经元的功能被抑制,而异常行为则再次出现;最后随着配体逐渐被代谢,转分化神经元的功能又得到恢复,而异常行为也同步改善。由于这些化学遗传学受体只表达在转分化神经元上,而与其他细胞无关,所以该实验能有力证明动物行为改善是直接来自于新生的神经元。
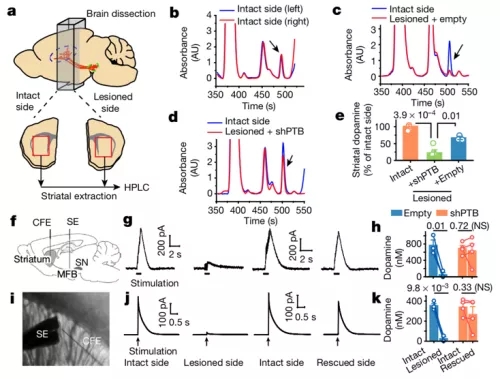
对于问题二,如何能实时地记录脑内的神经递质释放呢?当时,北京大学生命科学学院的 “探针王子” 李毓龙的 GRAB 探针系列还没有正式发表,要想记录中脑神经元投射到纹状体区域的神经递质多巴胺的释放量,比较成熟的实验技术是基于氧化还原原理的碳纤电极记录法。付向东的团队此前并没有使用这一实验方法的经验,于是向北大分子医学所的周专教授进行求助—— 周专团队昔日的成员、已经在泸州的西南医科大学建立实验室的康新江研究员此前关于记录多巴胺释放积累了丰富的经验。于是康新江临时回到在北京大学昔日工作的地方,在收到付向东从圣地亚哥寄来的小鼠后立刻开展实验:他用一根电极刺激黑质纹状体通路(预期激活那些由星形细胞转分化形成的神经元),同时用另一根记录电极在纹状体区域(转分化形成的新神经元投射到的脑区)进行记录。
在这个实验中,记录电极在完整的脑区内能记录到电刺激引起的多巴胺释放,而在因帕金森模型诱导发生损伤的脑区所记录到多巴胺释放信号会显著降低。如果在损伤的大脑中出现转分化神经元后,能记录到多巴胺信号的增强,就可以为转分化神经元具有神经信号传送功能提供强力的证据。
周专和付向东在这个合作中采取了双盲的实验设计:即康新江在记录电刺激诱导的多巴胺释放时,并不知道手中的样品是否损伤、是否表达了转分化神经元。康新江将数据整理后,与付向东手中的小鼠编号核对,这样可以尽可能地减少主观因素对实验结果的影响。
实验的结果十分清晰,完全符合实验前的预期。审稿人的第二个问题由此迎刃而解。
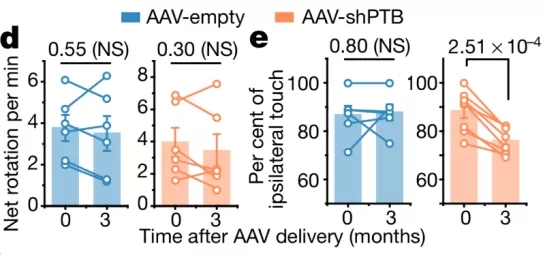
第三个问题回答起来并不困难,只要在年老的小鼠中进行类似的实验处理即可—— 钱浩此前的转分化实验主要使用的是两个月大的小鼠,现在将其应用到年龄为一年大的小鼠身上。但这一回,实验结果却有点令人费解。
为了检验转分化神经元对小鼠运动能力的恢复作用,钱浩使用了两种不同的行为学实验:此前在两个月大的小鼠中转分化神经元在两种行为学实验都能产生显著的修复效果;但在一年大的小鼠中,两个行为学实验却只有一个表现出修复效果,另一个和对照组没有显著的差异。
“对于这个结果的解释需要格外谨慎,” 付向东表示,“我们认为这个结果表明转分化神经元在一年大的帕金森模型小鼠的运动能力修复中能取得 ‘部分成功’(partial success),但目前我们还不能得出更明确的结论—— 这是一个既不能说 yes 也不能说 no 的实验结果。”
钱浩和付向东投入更多的时间精力去确认这一结果的可重复性,并在重新投回到《自然》的稿件中将这部分数据附在了 “补充图表”(extended figure)中,并表示这部分数据由于比较模糊,可能考虑会从论文中去除。
付向东与周专合作开展的激活-多巴胺释放记录实验[9]
化学遗传学实验论证转分化形成的新神经元直接导致运动能力的修复[9]
年老小鼠行为学实验结果[9]
再次看到经过一年多时间打磨的论文后,审稿人们深深被论文扎实的论证折服,一致同意发表。其中一位审稿人还强烈建议付向东将年老小鼠的实验结果放在主图表(main figure)中—— 在他看来,这个结果非常有说服力。作为帕金森模型的专家,他认为在该行为学实验中无显著差异,而在另一个行为学实验中表现出显著修复效果,正是他所意料中的结果。因为有些行为学模型受年龄的影响,进而进一步支持付向东论文的结论,即通过敲低 PTB 诱导产生转分化神经元在年老帕金森模型鼠中也能达到一定程度的修复效果。
正当万事俱备,编辑准备正式接受付向东的文章之时,《细胞》在2020年4月8日在线发表了中科院神科所杨辉研究员利用 CRISPR 技术敲低 PTB 治疗视网膜疾病的论文。[14]
看到自己的工作被同行 “抢发”,付向东不禁心中一凉。但很快《自然》的编辑为他吃下了一颗定心丸:原来,在2019年1月的时候,杨辉团队的这篇论文首先投到了《自然》,并且被编辑交给了同一批审稿人进行同行评议。但几位审稿人都认为杨辉团队的论文尽管与付向东的论文有异曲同工之妙,但在数据的扎实程度方面略差一筹,最终将这篇论文拒绝。
“《自然》的编辑告诉我们,他们坚信在为发现优先权盖棺定论时,论文提交的时间要比实际发表的时间更重要。而《自然》作为一家有着百年传统的顶级科学期刊,与其抢在其他期刊之前发表重要的原创发现,更在意支撑论文结论的证据的可靠性。”
2020年6月25日,付向东的文章作为封面文章在《自然》亮相,在国内外学术界均引起了很大的关注。付向东表示,“我计划将剩余的科学生涯,完全投入到将分子生物学应用到神经退行性疾病的治疗研究中。”
参考文献
1. Boutz, P. L. et al. A post-transcriptional regulatory switch in polypyrimidine tract-binding proteins reprograms alternative splicing in developing neurons. Genes & development 21, 1636-1652 (2007).
2. Coutinho-Mansfield, G. C., Xue, Y., Zhang, Y. & Fu, X.-D. PTB/nPTB switch: a post-transcriptional mechanism for programming neuronal differentiation. Genes & development 21, 1573-1577 (2007).
3. Ule, J. et al. CLIP identifies Nova-regulated RNA networks in the brain. Science 302, 1212-1215 (2003).
4. Garcia-Blanco, M. A., Jamison, S. F. & Sharp, P. A. Identification and purification of a 62,000-dalton protein that binds specifically to the polypyrimidine tract of introns. Genes & development 3, 1874-1886 (1989).
5. Singh, R., Valcarcel, J. & Green, M. R. Distinct binding specificities and functions of higher eukaryotic polypyrimidine tract-binding proteins. Science 268, 1173-1176 (1995).
6. Hu, J., Qian, H., Xue, Y. & Fu, X.-D. PTB/nPTB: master regulators of neuronal fate in mammals. Biophysics reports 4, 204-214 (2018).
7. Xue, Y. et al. Genome-wide analysis of PTB-RNA interactions reveals a strategy used by the general splicing repressor to modulate exon inclusion or skipping. Molecular cell 36, 996-1006 (2009).
8. Xue, Y. et al. Direct conversion of fibroblasts to neurons by reprogramming PTB-regulated microRNA circuits. Cell 152, 82-96 (2013).
9. Qian, H. et al. Reversing a model of Parkinson’s disease with in situ converted nigral neurons. Nature 582, 550-556 (2020).
10. Xue, Y. et al. Sequential regulatory loops as key gatekeepers for neuronal reprogramming in human cells. Nature neuroscience 19, 807-815 (2016).
11. di Val Cervo, P. R. et al. Induction of functional dopamine neurons from human astrocytes in vitro and mouse astrocytes in a Parkinson's disease model. Nature biotechnology 35, 444-452 (2017).
12. Barker, R. A., Götz, M. & Parmar, M. New approaches for brain repair—from rescue to reprogramming. Nature 557, 329-334 (2018).
13. Cohen, D. E. & Melton, D. Turning straw into gold: directing cell fate for regenerative medicine. Nature Reviews Genetics 12, 243-252 (2011).
14. Zhou, H. et al. Glia-to-Neuron Conversion by CRISPR-CasRx Alleviates Symptoms of Neurological Disease in Mice. Cell (2020).

扫描上面二维码在移动端打开阅读