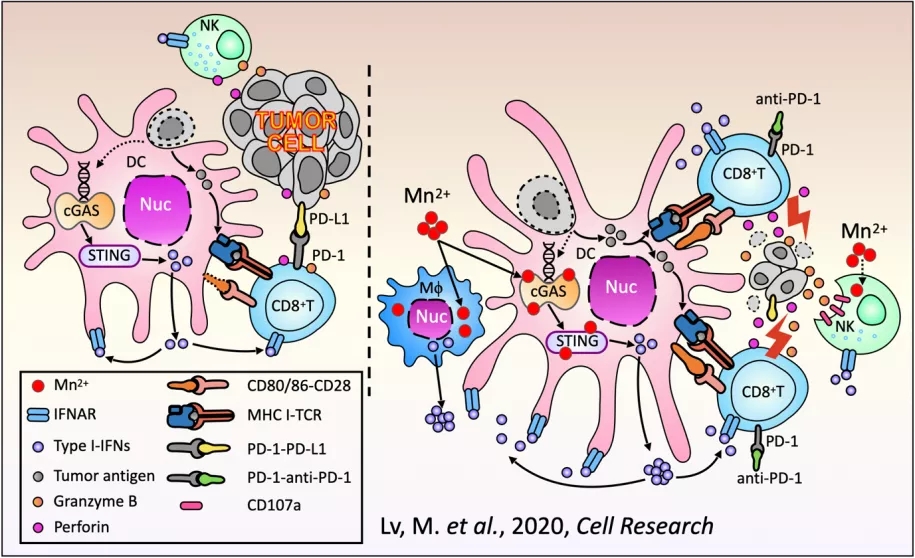
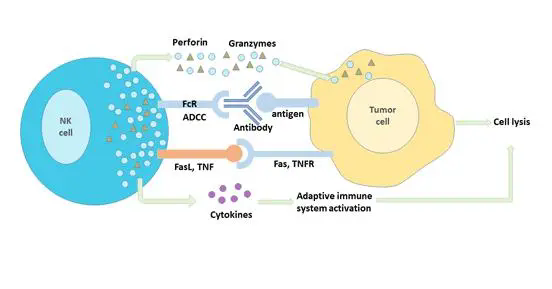
天然免疫对肿瘤的免疫监视至关重要,如自然杀伤细胞(NK)可以不依赖抗体和补体直接杀伤肿瘤细胞【1】,巨噬细胞(Mφ)和树突状细胞(DC)负责发现肿瘤细胞并将肿瘤抗原递呈给并激活细胞毒性T细胞(CD8+T细胞)【2】,后者特异性识别、杀灭肿瘤细胞。肿瘤细胞则进化出了一系列逃逸机制,以躲避宿主免疫系统的攻击【3】。其中以上调免疫检查点分子的表达最为著名,如肿瘤细胞高表达PD-1和/或CTLA-4抑制T细胞的活化。PD-1和CTLA-4抗体的临床应用是当今最令人激动的突破和国际公认最有前景的癌症治愈手段,获得了2018年的诺贝尔生理学或医学奖【4-10】。然而,免疫检查点抑制剂只对大约20%的肿瘤患者有效【11-13】,如何提高肿瘤患者对免疫治疗的反应性是亟待解决的关键问题。近期一系列的研究发现,肿瘤组织中(旁)Mφ和DC内感受胞质DNA的cGAS-STING通路可以被肿瘤细胞来源的DNA激活,促进这些抗原递呈细胞的成熟,进而将肿瘤特异性抗原递呈给T细胞,对于复苏机体的抗肿瘤免疫反应起到至关重要的作用【14-20】。目前全球许多科研机构和制药公司纷纷投入巨资,寻找cGAS-STING通路的激动剂【21-25】。然而目前已知的这些激动剂都制备复杂,成本高昂,而且无法有效进入细胞,因此治疗效果不理想【26,27】。北京大学生科院蒋争凡实验室是世界上同期发现STING分子与功能的实验室之一(命名为ERIS,武汉大学舒红兵实验室命名其为MITA)【28-31】,近年来在该领域取得了一系列成果。2018年,蒋争凡实验室发表在Immunity的工作发现锰离子(Mn2+)在激活cGAS-STING通路中的重要作用:病毒感染细胞导致线粒体、高尔基体等贮锰细胞器中的Mn2+释放到胞质及胞外。胞质中显著升高的Mn2+浓度(约60倍)对cGAS-STING通路具有双重激活的作用:Mn2+能够上万倍地提升cGAS对于DNA的检测灵敏度并促进其合成第二信使cGAMP;Mn2+上百倍地增强STING与cGAMP的结合能力。这样,释放到胞质的Mn2+使得细胞的cGAS-STING通路处于超活化的状态,极大地提升其对胞质DNA的响应能力,甚至使其在原本不具激活能力的DNA水平下也能够被激活。而当小鼠缺乏锰元素时,其抵抗DNA病毒的能力受到显著抑制(详见BioArt报道:蒋争凡组Immunity揭示锰离子抵御DNA病毒入侵的新机制)【32】。2020年8月18日,蒋争凡实验室与苏晓东实验室合作发表在Cell Reports的工作进一步阐释了Mn2+激活cGAS的分子机制,发现Mn2+还可以直接激活cGAS(完全不依赖dsDNA),而且Mn2+催化与Mg2++dsDNA催化cGAS合成cGAMP的过程显著不同【33】。Mn2+-cGAS结合导致cGAS构象发生类似于DNA-cGAS结合的变化,但催化中心的结构明显不同,导致cGAMP在cGAS酶催化中心的合成过程也发生很大变化。这个研究进一步证明Mn2+DNA以外的第二个cGAS激活剂。然而,锰元素/锰离子(Mn2+)在抗肿瘤免疫中的作用尚不清楚。2020年8月24日,蒋争凡实验室与解放军总医院第一医学中心生物治疗科韩为东团队合作在Cell Research上以Research Article的形式在线发表了肿瘤免疫领域的最新成果“Manganese is critical for antitumor immune responses via cGAS-STING and improves the efficacy of clinical immunotherapy”。在本研究中【34】,蒋争凡实验室首先发现缺锰小鼠体内的肿瘤细胞生长显著加快、肿瘤肺转移显著增多,说明在生理条件下,作为必需微量元素的锰在肿瘤免疫中也发挥必不可少的作用。
1. Kumar,V. & McNerney, M. E. A new self: MHC-class-I-independentnatural-killer-cell self-tolerance. NatRev Immunol 5, 363-374,doi:10.1038/nri1603 (2005).2. Trombetta, E. S. & Mellman, I. Cellbiology of antigen processing in vitro and in vivo. Annu Rev Immunol 23,975-1028, doi:10.1146/annurev.immunol.22.012703.104538 (2005).3. O'Donnell, J. S., Teng, M. W. L. & Smyth,M. J. Cancer immunoediting and resistance to T cell-based immunotherapy. Nat Rev Clin Oncol 16, 151-167, doi:10.1038/s41571-018-0142-8 (2019).4. Brunet, J. F. et al. A new member of the immunoglobulin superfamily--CTLA-4. Nature 328, 267-270, doi:10.1038/328267a0 (1987).5. Walunas, T. L. et al. CTLA-4 can function as a negative regulator of T cellactivation. Immunity 1, 405-413,doi:10.1016/1074-7613(94)90071-x (1994).6. Waterhouse, P. et al. Lymphoproliferative disorders with early lethality in micedeficient in Ctla-4. Science 270, 985-988, doi:10.1126/science.270.5238.985(1995).7. Tivol, E. A. et al. Loss of CTLA-4 leads to massive lymphoproliferation andfatal multiorgan tissue destruction, revealing a critical negative regulatoryrole of CTLA-4. Immunity 3, 541-547, doi:10.1016/1074-7613(95)90125-6(1995).8. Leach, D. R., Krummel, M. F. & Allison,J. P. Enhancement of antitumor immunity by CTLA-4 blockade. Science 271, 1734-1736, doi:10.1126/science.271.5256.1734 (1996).9. Nishimura, H., Nose, M., Hiai, H., Minato, N.& Honjo, T. Development of lupus-like autoimmune diseases by disruption ofthe PD-1 gene encoding an ITIM motif-carrying immunoreceptor. Immunity 11, 141-151, doi:10.1016/s1074-7613(00)80089-8 (1999).10 Ishida, Y., Agata, Y., Shibahara, K. &Honjo, T. Induced expression of PD-1, a novel member of the immunoglobulin genesuperfamily, upon programmed cell death. EMBOJ 11, 3887-3895 (1992).11 Topalian, S. L., Drake, C. G. & Pardoll, D.M. Immune checkpoint blockade: a common denominator approach to cancer therapy.Cancer Cell 27, 450-461, doi:10.1016/j.ccell.2015.03.001 (2015).12 Huang, A. C.et al. T-cell invigoration to tumour burden ratio associated with anti-PD-1response. Nature 545, 60-65, doi:10.1038/nature22079 (2017).13 Ribas, A.et al. Association of Pembrolizumab With Tumor Response and Survival AmongPatients With Advanced Melanoma. JAMA315, 1600-1609,doi:10.1001/jama.2016.4059 (2016).14 Deng, L.et al. STING-Dependent Cytosolic DNA Sensing Promotes Radiation-InducedType I Interferon-Dependent Antitumor Immunity in Immunogenic Tumors. Immunity 41, 843-852, doi:10.1016/j.immuni.2014.10.019 (2014).15 Woo, S. R.et al. STING-dependent cytosolic DNA sensing mediates innate immunerecognition of immunogenic tumors. Immunity41, 830-842,doi:10.1016/j.immuni.2014.10.017 (2014).16 Woo, S. R., Corrales, L. & Gajewski, T. F.The STING pathway and the T cell-inflamed tumor microenvironment. Trends Immunol 36, 250-256, doi:10.1016/j.it.2015.02.003 (2015).17 Chen, Q., Sun, L. & Chen, Z. J. Regulationand function of the cGAS-STING pathway of cytosolic DNA sensing. Nat Immunol 17, 1142-1149, doi:10.1038/ni.3558 (2016).18 Wang, H.et al. cGAS is essential for the antitumor effect of immune checkpointblockade. Proc Natl Acad Sci U S A 114, 1637-1642,doi:10.1073/pnas.1621363114 (2017).19 Dunn, G. P.et al. A critical function for type I interferons in cancer immunoediting. Nat Immunol 6, 722-729, doi:10.1038/ni1213 (2005).20 Dubensky, T. W., Jr. & Reed, S. G.Adjuvants for cancer vaccines. SeminImmunol 22, 155-161,doi:10.1016/j.smim.2010.04.007 (2010).21 Roberts, Z. J., Ching, L. M. & Vogel, S. N.IFN-beta-dependent inhibition of tumor growth by the vascular disrupting agent5,6-dimethylxanthenone-4-acetic acid (DMXAA). J Interferon Cytokine Res 28,133-139, doi:10.1089/jir.2007.0992 (2008).22 Conlon, J.et al. Mouse, but not human STING, binds and signals in response to thevascular disrupting agent 5,6-dimethylxanthenone-4-acetic acid. J Immunol 190, 5216-5225, doi:10.4049/jimmunol.1300097 (2013).23 Corrales, L.et al. Direct Activation of STING in the Tumor Microenvironment Leads toPotent and Systemic Tumor Regression and Immunity. Cell Rep 11, 1018-1030,doi:10.1016/j.celrep.2015.04.031 (2015).24 Demaria, O.et al. STING activation of tumor endothelial cells initiates spontaneousand therapeutic antitumor immunity. ProcNatl Acad Sci U S A 112,15408-15413, doi:10.1073/pnas.1512832112 (2015).25 Ramanjulu, J. M. et al. Design of amidobenzimidazole STING receptor agonists withsystemic activity. Nature 564, 439-443,doi:10.1038/s41586-018-0705-y (2018).26 Lara, P. N., Jr. et al. Randomized phase III placebo-controlled trial ofcarboplatin and paclitaxel with or without the vascular disrupting agentvadimezan (ASA404) in advanced non-small-cell lung cancer. J Clin Oncol 29,2965-2971, doi:10.1200/JCO.2011.35.0660 (2011).27 Bhattacharya, N. et al. Normalizing Microbiota-Induced Retinoic Acid DeficiencyStimulates Protective CD8(+) T Cell-Mediated Immunity in Colorectal Cancer. Immunity 45, 641-655, doi:10.1016/j.immuni.2016.08.008 (2016).28 Ishikawa, H. & Barber, G. N. STING is anendoplasmic reticulum adaptor that facilitates innate immune signalling. Nature 455, 674-678, doi:10.1038/nature07317 (2008).29 Zhong, B.et al. The adaptor protein MITA links virus-sensing receptors to IRF3transcription factor activation. Immunity29, 538-550,doi:10.1016/j.immuni.2008.09.003 (2008).30 Sun, W.et al. ERIS, an endoplasmic reticulum IFN stimulator, activates innateimmune signaling through dimerization. ProcNatl Acad Sci U S A 106,8653-8658, doi:10.1073/pnas.0900850106 (2009).31 Jin, L.et al. MPYS, a novel membrane tetraspanner, is associated with majorhistocompatibility complex class II and mediates transduction of apoptoticsignals. Mol Cell Biol 28, 5014-5026, doi:10.1128/MCB.00640-08(2008).32 Wang, C.et al. Manganese Increases the Sensitivity of the cGAS-STING Pathway forDouble-Stranded DNA and Is Required for the Host Defense against DNA Viruses. Immunity 48, 675-687 e677, doi:10.1016/j.immuni.2018.03.017 (2018).33 Zhao, Z. M., Z.; Wang, B.; Guan, Y.; Su, X.;Jiang, Z. Mn2+ Directly Activates cGAS and Triggers A DistinctCatalytic Synthesis of 2’3’-cGAMP.CellReports Accepted (2020).34 Lv, M. etal. Manganese is critical for antitumor immune responses via cGAS-STING andimproves the efficacy of clinical immunotherapy. Cell Res Accpeted(2020).