摘要
前言


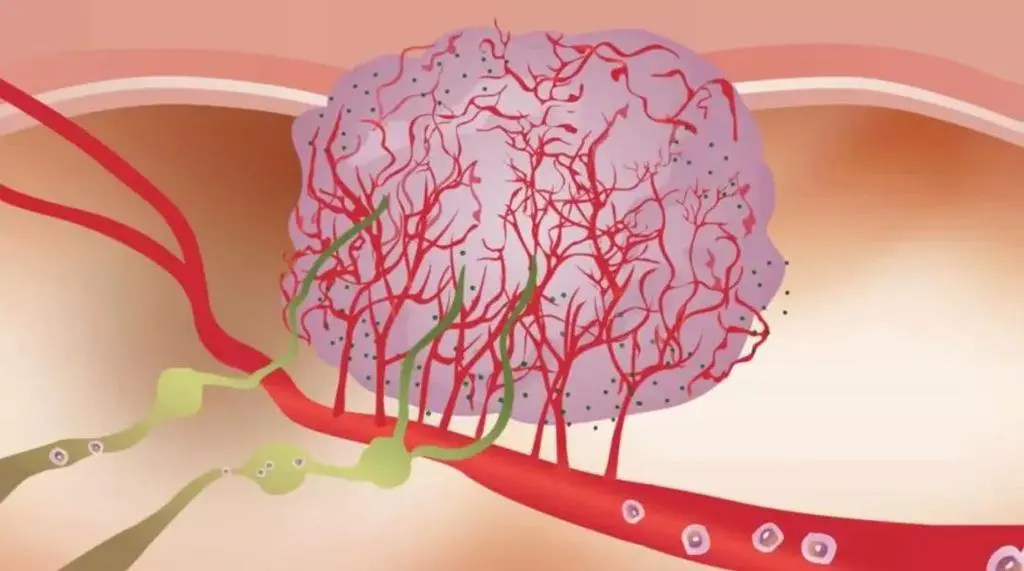
图1:TME中的癌细胞代谢和异常
癌细胞具有上调葡萄糖的糖酵解分解代谢,形成乳酸盐的能力,即使在氧气充足的条件下(有氧糖酵解),该效应被称为“ Warburg效应”,也被认为是恶性肿瘤的标志。现在人们越来越清楚地认识到,癌症的代谢也具有异质性,癌细胞可以参与各种代谢程序以满足生长和增殖的需求,除有氧糖酵解之外,线粒体呼吸也至关重要。我们可以发现具有高代谢活性的癌细胞(图1)会对TME产生深远的影响,导致营养物质消耗,缺氧,酸性和代谢产物的产生,这些代谢产物在浓度过高下可能具有毒性。来自TME的大量葡萄糖通过有氧糖酵解代谢,产生大量的乳酸和H +,从而降低了肿瘤内的pH。也就是说,产生乳酸的糖酵解和氧化磷酸化(OXPHOS)之间的平衡可能取决于缺氧的程度,缺氧的程度在TME中既可以是异质的,又可以是广泛的。值得注意的是,在中度低氧区域,线粒体呼吸产生的CO2被细胞外碳酸酐酶水合,形成HCO3-和H +。因此,氧化代谢可能是TME中细胞外酸化的重要且易被忽视的来源。
肿瘤免疫微环境

表 1 肿瘤微环境中的细胞组成
抗肿瘤反应的代谢
抗肿瘤效应T细胞的葡萄糖代谢
TME中的T细胞和葡萄糖受限
氨基酸与抗肿瘤T细胞反应
脂质代谢与T细胞
免疫记忆的代谢
缺氧与抗肿瘤T细胞反应
毒性代谢物

图2:TME中的代谢紊乱抑制T细胞功能
肿瘤中特定氨基酸的积累也可以抑制Teff细胞反应。在这方面,研究得最好的是色氨酸代谢产物,尤其是犬尿氨酸,其作用是通过IDO1的活性产生。与对照相比,犬尿氨酸在体外引起活化CD8 + T细胞上PD1协同抑制途径的上调。与对照相比,在用外源性犬尿氨酸处理的小鼠模型中,在肿瘤浸润的CD8 + T细胞上也观察到PD1的这种上调。来自经犬尿氨酸处理的荷瘤小鼠的肿瘤浸润CD8 + T细胞产生的IFNγ和TNF较少。
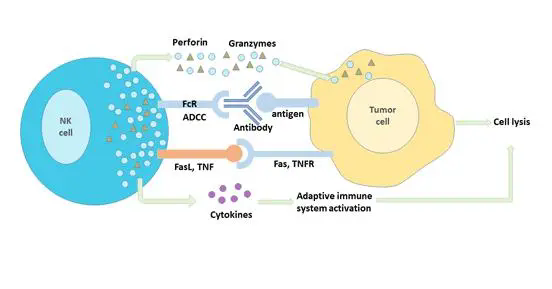
代谢和天然效应反应
癌症免疫逃逸的新陈代谢
适应性免疫抑制的代谢
天然免疫抑制的代谢
利用不同代谢的可塑性
免疫检查点封闭与免疫代谢
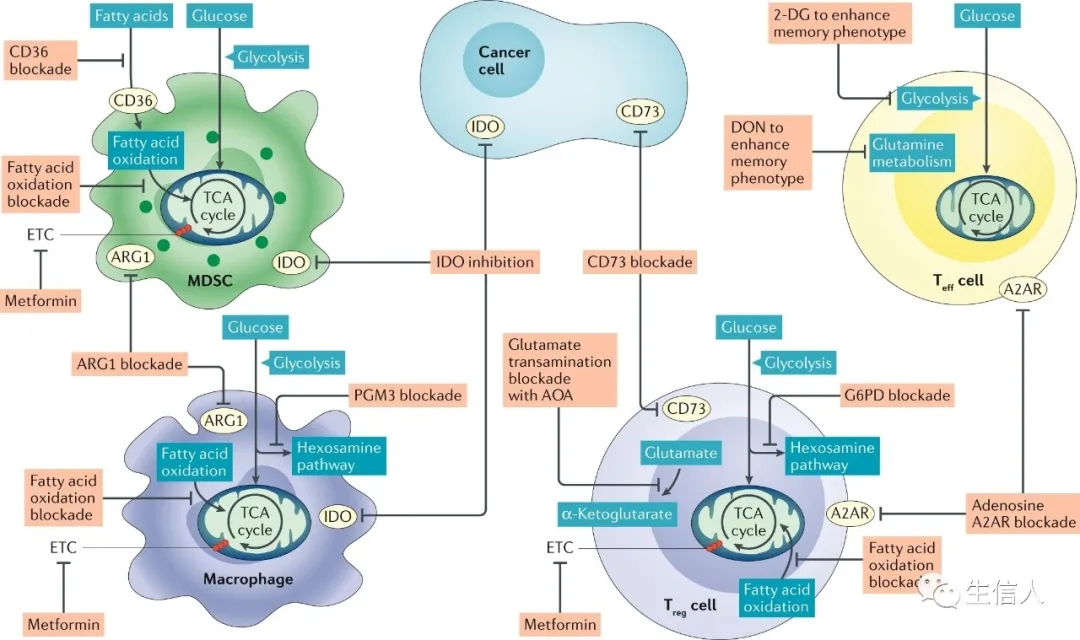
图3:增强癌症免疫反应的潜在代谢靶点
结论与展望
扫描上面二维码在移动端打开阅读