

免疫!作为科研探究的一大领域,广受关注,尤其常与肿瘤挂钩,免疫治疗也是科研人的探索热点。但免疫这块“大饼”,可不是那么容易“啃”的喔~本期就先带小伙伴们了解下常见的免疫细胞与免疫分子!
T 细胞如何被活化?
当 T 细胞在受感染细胞或吞噬细胞表面遇到与 MHC 分子偶联的同源抗原时,T 淋巴细胞活化途径就会被触发。
T 细胞是许多免疫疗法的靶点,如检查点抑制剂、双特异性 T 细胞接合剂和免疫激动剂等。同时,T 细胞本身也已被研究用于治疗血液系统恶性肿瘤,方法是使用嵌合抗原受体添加 (CAR addition) 或自体过继转移 (过继转移策略包括体外扩增和激活自体肿瘤反应性 CTL,然后再回输给患者)。
在实体瘤中,尽管针对 PD-1、CTLA-4 和 LAG3 的检查点抑制剂取得了一些成功,但大多数方法并未提高总体生存率。此外,激活 T 细胞杀死癌症的方法可能会诱发严重的免疫相关不良事件,在某些情况下甚至会导致死亡[2]。

初始 B 细胞 (Naïve B cell) 响应激活信号增殖并分化为效应细胞。在早期的多克隆反应中,可以产生分泌多反应性抗体的短寿命浆细胞 (Short-lived plasma cell)。调节性 B 细胞 (Regulatory B cell) 也可以通过分泌调节 T 细胞应答的 IL-10、IL-17、IL-35 和 TGFβ 来诱导和发挥免疫抑制功能。
持续的 B 细胞激活导致有组织的淋巴样结构进一步分化和选择,称为生发中心(Germinal centers, GCs)。NF-κB 的激活和激活诱导胞苷脱氨酶 (AID) 的上调,通过抗体重链的体细胞高突变和类转换重组诱导抗体亲和成熟。这最终导致特异性的、长寿命的浆细胞 (Long-lived plasma cell) 和记忆 B 细胞的分化,从而赋予保护性免疫。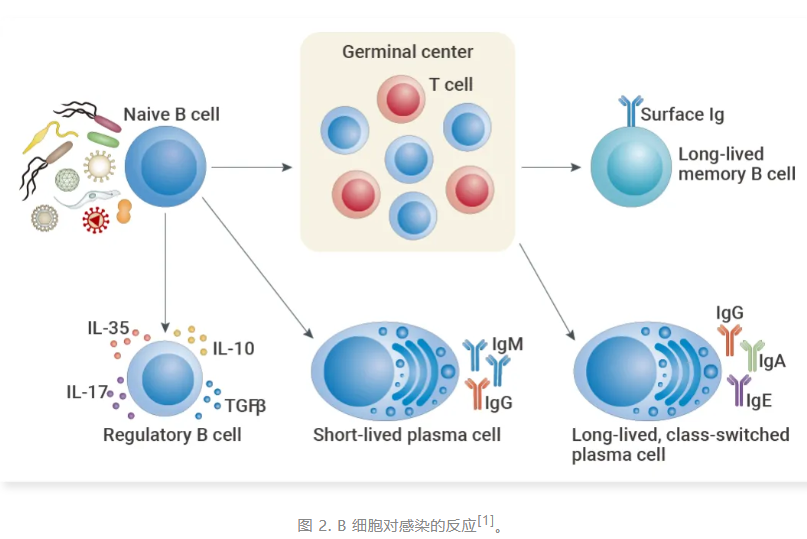

正常情况下,免疫系统会将主要组织相容性复合体 (MHC) 分子(特别是人类白细胞抗原 (HLA))中呈递的自身肽识别为“安全”的,因此将其忽略。这被称为自我耐受。
在“缺失自我”模型中,当 NK 细胞的抑制性受体因“自我识别分子”(HLA) 分子组不完整或不兼容而无法识别靶细胞时,NK 细胞就会被激活[4]。 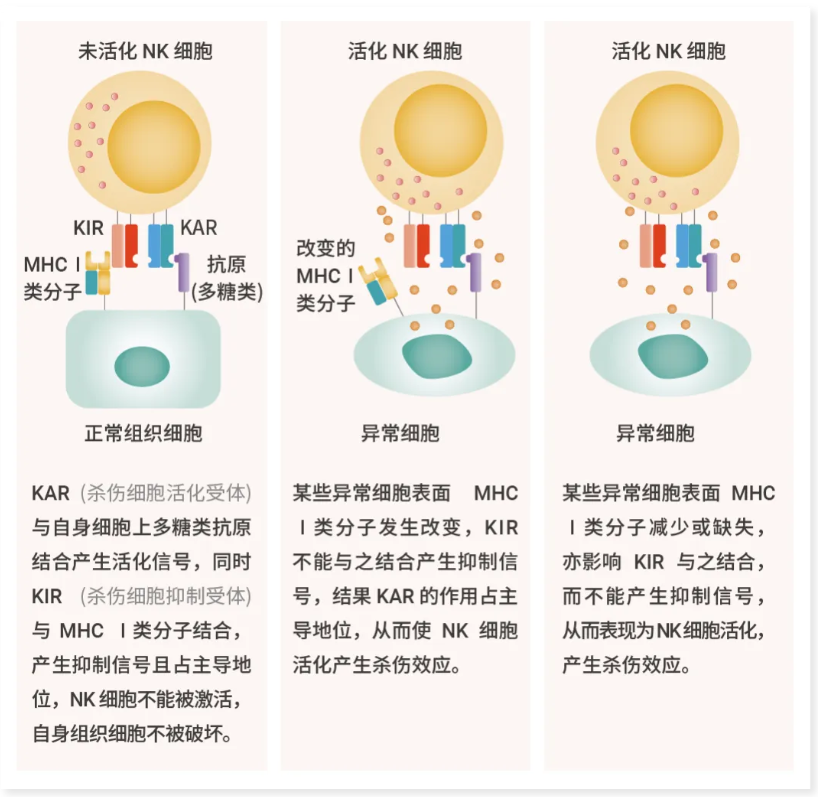
图 3. NK 细胞 KIR 和 KAR 的作用。
在癌症中,受感染的、外来的或癌细胞会将正常的自身肽交换为“非自身”肽,从而使免疫系统能够靶向该肽。然而,肿瘤细胞常常会通过下调 MHC-I 分子的表面表达来逃避这种免疫监视,从而绕过 CD8+ T 细胞杀伤。为了应对这种情况,较低的 MHC 表达会释放宿主 NK 细胞抑制受体并激活它们,从而杀死癌细胞[4]。 
当巨噬细胞摄取病原体时,首先透过吞噬作用摄取病原体 (a),形成一吞噬体。随后后溶酶体融入吞噬体并形成吞解体;病原体被酶所分解 (b)。接着 (c) 废料被排出或同化 (后者则未被绘成图象)。

树突状细胞几乎存在于所有组织中,它们检测体内平衡失衡并处理抗原以呈递给 T 细胞,从而建立先天免疫反应和适应性免疫反应之间的联系。
树突状细胞有两种不同的功能状态:"成熟" (Mature) 和 "未成熟" (IMmature)。成熟树突状细胞的标志是能够激活次级淋巴器官中的抗原特异性初始 T 细胞。成熟会启动代谢、细胞和基因转录程序,使树突状细胞从外周组织迁移到次级淋巴器官中的 T 依赖性区域,T 淋巴细胞激活抗原呈递可能在此发生[5]。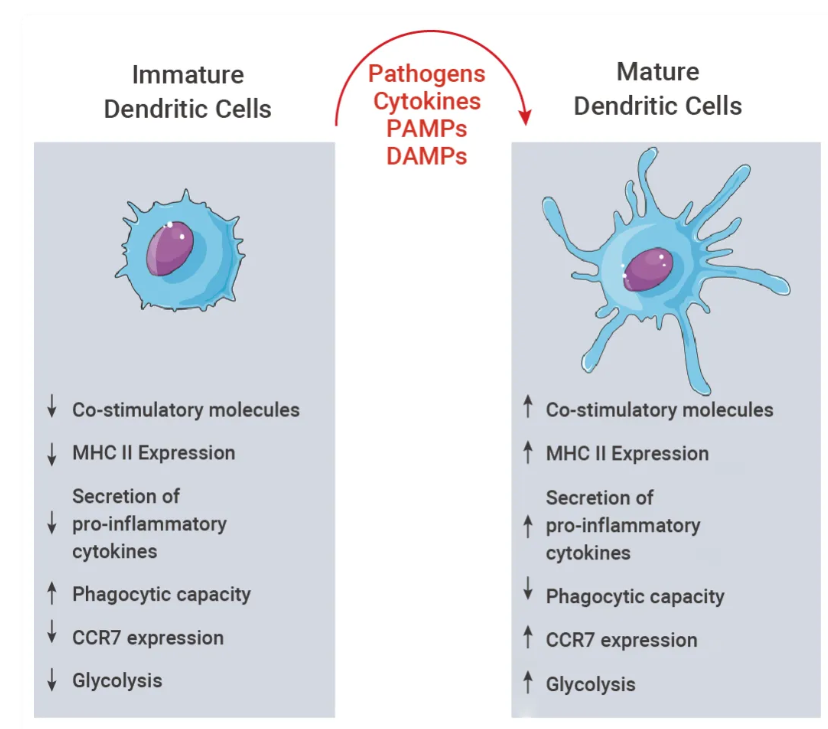
图 5. 树突状细胞活化[5]。
细胞外信号(例如 PAMP 或 DAMP)会触发未成熟树突状细胞发生改变,最终导致表面蛋白、细胞内通路和代谢活动发生显著变化。
粒细胞 (Granulocyte) 用 Wright’s 染色可分辨出三种颗粒白细胞即中性粒细胞、嗜酸性粒细胞和嗜碱性粒细胞。
可分为“专业” (Professional) 抗原呈递细胞和“非典型” (Atypical) 抗原呈递细胞。
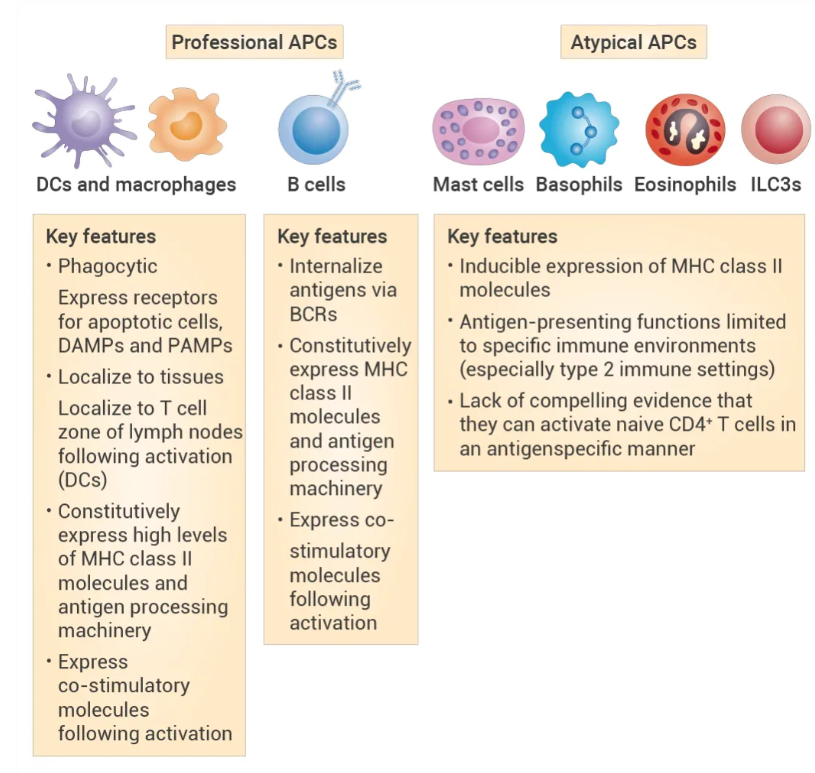
图 7. “专业”抗原提呈细胞和“非典型”抗原提呈细胞的关键抗原提呈功能[7]。
CD 即是位于细胞膜上的一类分化抗原的总称,CD 后面的序号代表一个或一类分化抗原分子。
CD 分子通常用作免疫抗原辨识的细胞标记,可借此观察细胞表面的分子并辨识该细胞。
例如:不同血细胞表面分布有不同的 CD 分子,由此可以对不同的血细胞进行鉴定并分离。
三个保守的 CAM 结构域包括胞内结构域 (与细胞骨架相互作用)、跨膜结构域和胞外结构域。 
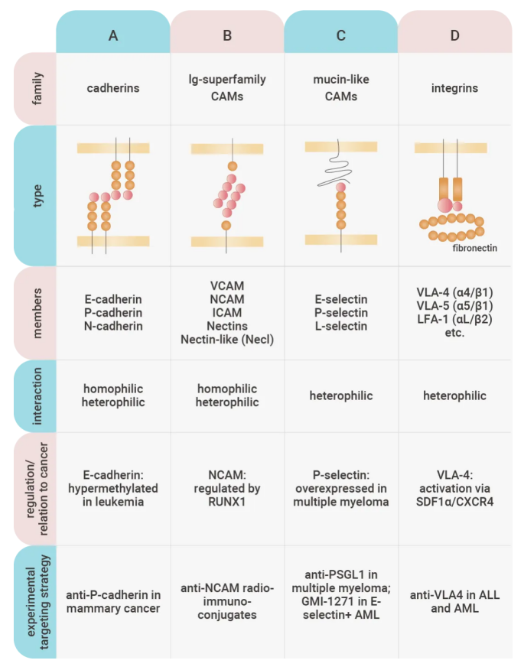
图 8. 四种主要细胞粘附分子家族的结构和功能特性[8]。

细胞因子受体 (Cytokine receptor): 是指能与细胞因子相结合的受体。
免疫系统的细胞通过交换分泌的细胞因子进行交流。根据消耗细胞因子的细胞的空间位置和身份,这种交流可以是自分泌 (向自身发出信号)、旁分泌 (向邻近细胞发出信号) 或内分泌 (通过循环向全局发出信号)。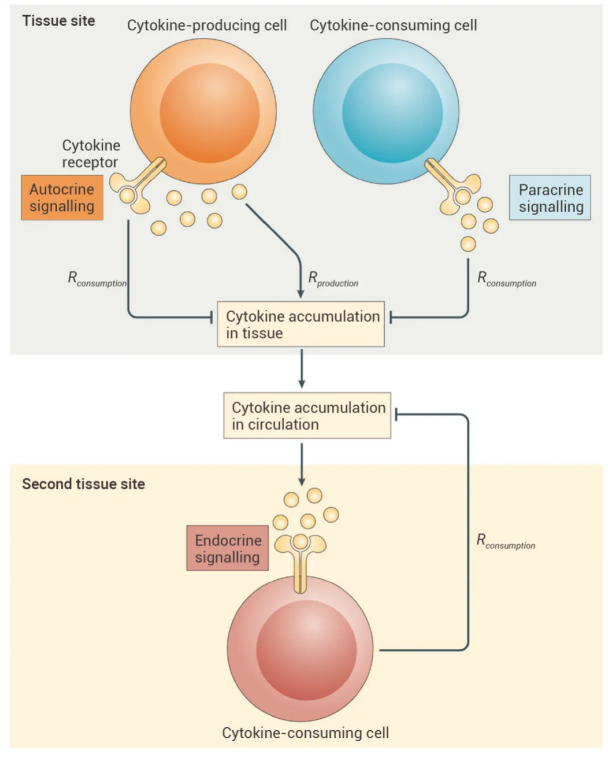
图 9. 细胞因子介导的细胞间通讯的三种模式[9]。
当受到几种触发剂中的一种刺激时,该系统中的蛋白酶会切割特定的蛋白质,释放细胞因子并启动进一步切割的级联放大反应。
补体系统激活或补体固定级联反应的最终结果是刺激吞噬细胞清除外来和受损物质,吸引更多的吞噬细胞参与炎症反应。
补体激活可通过经典、凝集素和替代三条独立途径实现。补体激活的关键步骤是 C3 裂解,其下游效应通过形成膜攻击复合物 (MAC) 导致细胞溶解和炎症信号传递。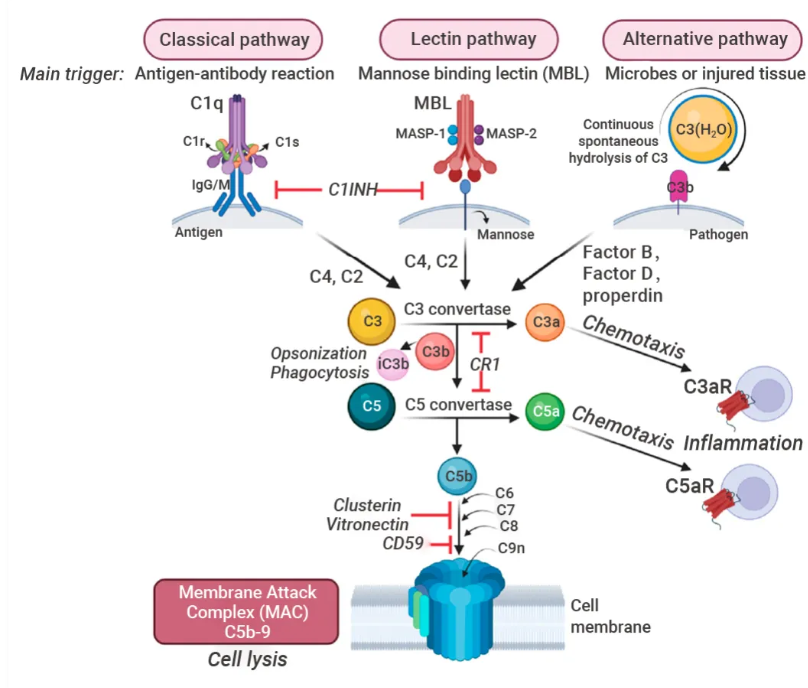
图 10. 补体级联[10]。
人类的 MHC 被称为 HLA (Human leukocyte antigen), 即人白细胞抗原。
抗体介导的天然蛋白识别不需要抗原处理,相比之下,T 淋巴细胞识别的蛋白质片段被抗原呈递细胞 (APC) 内的蛋白酶加工成短肽。抗原加工和呈递主要通过两种途径发生: MHC I 类途径和 MHC II 类途径。 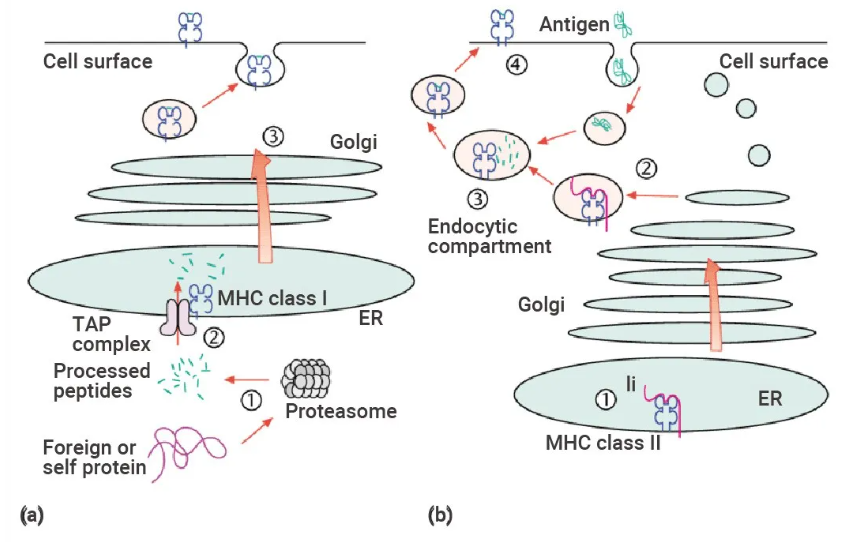
01 免疫细胞
1 T 淋巴细胞
T细胞,也称T淋巴细胞,淋巴细胞的主要组分,由胸腺内的淋巴干细胞分化而成,是淋巴细胞中数量最多,功能最复杂的一类细胞。
T 细胞如何被活化?
当 T 细胞在受感染细胞或吞噬细胞表面遇到与 MHC 分子偶联的同源抗原时,T 淋巴细胞活化途径就会被触发。
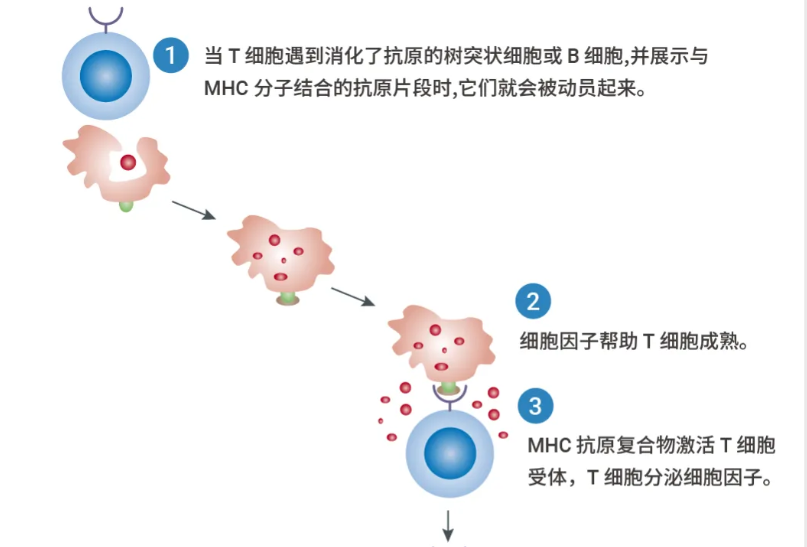
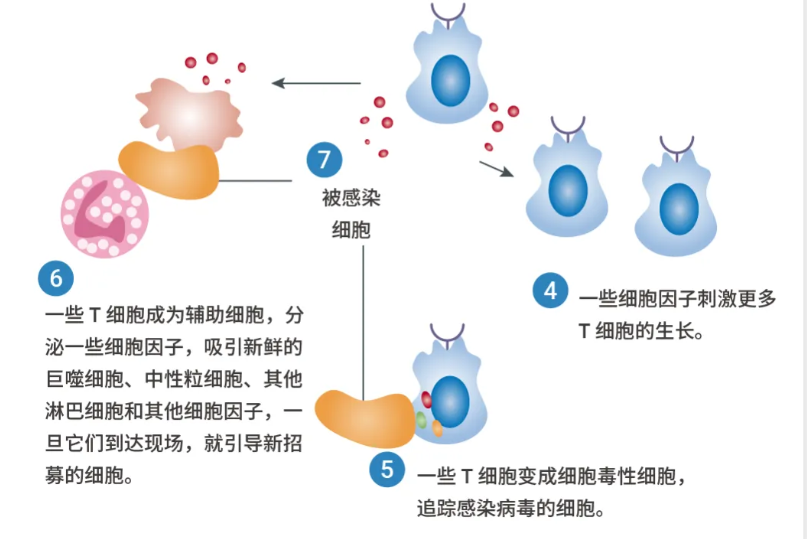
T 细胞是许多免疫疗法的靶点,如检查点抑制剂、双特异性 T 细胞接合剂和免疫激动剂等。同时,T 细胞本身也已被研究用于治疗血液系统恶性肿瘤,方法是使用嵌合抗原受体添加 (CAR addition) 或自体过继转移 (过继转移策略包括体外扩增和激活自体肿瘤反应性 CTL,然后再回输给患者)。
在实体瘤中,尽管针对 PD-1、CTLA-4 和 LAG3 的检查点抑制剂取得了一些成功,但大多数方法并未提高总体生存率。此外,激活 T 细胞杀死癌症的方法可能会诱发严重的免疫相关不良事件,在某些情况下甚至会导致死亡[2]。
2 B 淋巴细胞
B 细胞,也称 B 淋巴细胞,一种淋巴细胞的亚型,来源于骨髓中的造血干细胞,在骨髓中成熟。初始 B 细胞 (Naïve B cell) 响应激活信号增殖并分化为效应细胞。在早期的多克隆反应中,可以产生分泌多反应性抗体的短寿命浆细胞 (Short-lived plasma cell)。调节性 B 细胞 (Regulatory B cell) 也可以通过分泌调节 T 细胞应答的 IL-10、IL-17、IL-35 和 TGFβ 来诱导和发挥免疫抑制功能。
持续的 B 细胞激活导致有组织的淋巴样结构进一步分化和选择,称为生发中心(Germinal centers, GCs)。NF-κB 的激活和激活诱导胞苷脱氨酶 (AID) 的上调,通过抗体重链的体细胞高突变和类转换重组诱导抗体亲和成熟。这最终导致特异性的、长寿命的浆细胞 (Long-lived plasma cell) 和记忆 B 细胞的分化,从而赋予保护性免疫。
3 自然杀伤细胞
一种细胞质中具有大颗粒的淋巴细胞,20 世纪 70 年代首次被发现。NK 细胞具有杀伤功能,需要正常自体细胞发出抑制信号来防止不必要的细胞死亡。正常情况下,免疫系统会将主要组织相容性复合体 (MHC) 分子(特别是人类白细胞抗原 (HLA))中呈递的自身肽识别为“安全”的,因此将其忽略。这被称为自我耐受。
在“缺失自我”模型中,当 NK 细胞的抑制性受体因“自我识别分子”(HLA) 分子组不完整或不兼容而无法识别靶细胞时,NK 细胞就会被激活[4]。
图 3. NK 细胞 KIR 和 KAR 的作用。
在癌症中,受感染的、外来的或癌细胞会将正常的自身肽交换为“非自身”肽,从而使免疫系统能够靶向该肽。然而,肿瘤细胞常常会通过下调 MHC-I 分子的表面表达来逃避这种免疫监视,从而绕过 CD8+ T 细胞杀伤。为了应对这种情况,较低的 MHC 表达会释放宿主 NK 细胞抑制受体并激活它们,从而杀死癌细胞[4]。
4 巨噬细胞
单核细胞是人体免疫系统中的一种白细胞,产生于骨髓,在血管内为单核细胞,血管外就变成巨噬细胞。当巨噬细胞摄取病原体时,首先透过吞噬作用摄取病原体 (a),形成一吞噬体。随后后溶酶体融入吞噬体并形成吞解体;病原体被酶所分解 (b)。接着 (c) 废料被排出或同化 (后者则未被绘成图象)。
5 树突细胞
存在于哺乳动物的一种白细胞,是专业的抗原呈递细胞。树突状细胞几乎存在于所有组织中,它们检测体内平衡失衡并处理抗原以呈递给 T 细胞,从而建立先天免疫反应和适应性免疫反应之间的联系。
树突状细胞有两种不同的功能状态:"成熟" (Mature) 和 "未成熟" (IMmature)。成熟树突状细胞的标志是能够激活次级淋巴器官中的抗原特异性初始 T 细胞。成熟会启动代谢、细胞和基因转录程序,使树突状细胞从外周组织迁移到次级淋巴器官中的 T 依赖性区域,T 淋巴细胞激活抗原呈递可能在此发生[5]。
图 5. 树突状细胞活化[5]。
细胞外信号(例如 PAMP 或 DAMP)会触发未成熟树突状细胞发生改变,最终导致表面蛋白、细胞内通路和代谢活动发生显著变化。
6 粒细胞
一类细胞质中包含颗粒体的白细胞,绝大部分的粒细胞是中性粒细胞。
粒细胞 (Granulocyte) 用 Wright’s 染色可分辨出三种颗粒白细胞即中性粒细胞、嗜酸性粒细胞和嗜碱性粒细胞。
7 抗原呈递细胞
抗原呈递细胞 (Antigen-presenting cells, APC): 指能够摄取、加工处理抗原,并将处理过的抗原呈递给T细胞的一类免疫细胞。可分为“专业” (Professional) 抗原呈递细胞和“非典型” (Atypical) 抗原呈递细胞。

图 7. “专业”抗原提呈细胞和“非典型”抗原提呈细胞的关键抗原提呈功能[7]。
02 免疫分子
1 白细胞分化抗原
白细胞分化抗原 (Cluster of Differentiation,CD): 白细胞分化抗原,CD 分化簇或分化群,是细胞膜上的一类蛋白质或糖蛋白。指的是不同谱系的白细胞在正常分化成熟的不同阶段及活化过程中,出现或消失的细胞表面标记。CD 即是位于细胞膜上的一类分化抗原的总称,CD 后面的序号代表一个或一类分化抗原分子。

CD 分子通常用作免疫抗原辨识的细胞标记,可借此观察细胞表面的分子并辨识该细胞。
例如:不同血细胞表面分布有不同的 CD 分子,由此可以对不同的血细胞进行鉴定并分离。
表 1. 免疫表型的 CD 标记物。

2 黏附因子
细胞黏附分子 (Cell adhesion molecules, CAMs):位于细胞表面上的蛋白,是众多介导细胞间或细胞与细胞外基质间相互接触和结合分子的统称。三个保守的 CAM 结构域包括胞内结构域 (与细胞骨架相互作用)、跨膜结构域和胞外结构域。
图 8. 四种主要细胞粘附分子家族的结构和功能特性[8]。
3 免疫球蛋白超家族
免疫球蛋白超家族 (Immunoglobulin superfamily, IgSF):代表一大组蛋白质,其共同特征是它们都含有一个或多个细胞外 Ig 结构域,体内所有细胞均表达此类分子。4 细胞因子及其受体
细胞因子 (Cytokine):由免疫细胞 (如单核、巨噬细胞、T 细胞、B 细胞、NK 细胞等) 和某些非免疫细胞 (内皮细胞、表皮细胞、纤维母细胞等) 经刺激而合成、分泌的一类具有广泛生物学活性的小分子蛋白质。细胞因子受体 (Cytokine receptor): 是指能与细胞因子相结合的受体。

免疫系统的细胞通过交换分泌的细胞因子进行交流。根据消耗细胞因子的细胞的空间位置和身份,这种交流可以是自分泌 (向自身发出信号)、旁分泌 (向邻近细胞发出信号) 或内分泌 (通过循环向全局发出信号)。
图 9. 细胞因子介导的细胞间通讯的三种模式[9]。
5 补体
补体系统 (Complement system):又称补体级联反应,由一系列的蛋白质组成。当受到几种触发剂中的一种刺激时,该系统中的蛋白酶会切割特定的蛋白质,释放细胞因子并启动进一步切割的级联放大反应。
补体系统激活或补体固定级联反应的最终结果是刺激吞噬细胞清除外来和受损物质,吸引更多的吞噬细胞参与炎症反应。

补体激活可通过经典、凝集素和替代三条独立途径实现。补体激活的关键步骤是 C3 裂解,其下游效应通过形成膜攻击复合物 (MAC) 导致细胞溶解和炎症信号传递。
图 10. 补体级联[10]。
6 主要组织相容性复合体
主要组织相容性复合体 (Major histocompatibility complex, MHC):一组编码动物主要组织相容性抗原的基因群的统称。人类的 MHC 被称为 HLA (Human leukocyte antigen), 即人白细胞抗原。

抗体介导的天然蛋白识别不需要抗原处理,相比之下,T 淋巴细胞识别的蛋白质片段被抗原呈递细胞 (APC) 内的蛋白酶加工成短肽。抗原加工和呈递主要通过两种途径发生: MHC I 类途径和 MHC II 类途径。
图 11. 主要组织相容性复合体 (MHC) I 类和 II 类抗原加工和递呈途径的示意图[11]。
(a) 由蛋白酶体在细胞质中产生的肽类 ① 通过与抗原加工相关的肽转运体 (TAP) ② 转运到内质网 (ER),并装载到 MHC 类 I 分子上。随后,类 I 复合物通过高尔基体复合体 ③ 被运输到细胞表面。(b) MHC 类 II 分子与内质网中的不变链 (Ii) 相关 ①。Ii-相关复合物被靶向到内吞泡腔 ②,在那里从内吞蛋白产生的肽类被装载到 MHC 类 II 分子上 ③,并转移到细胞表面 ④。
(a) 由蛋白酶体在细胞质中产生的肽类 ① 通过与抗原加工相关的肽转运体 (TAP) ② 转运到内质网 (ER),并装载到 MHC 类 I 分子上。随后,类 I 复合物通过高尔基体复合体 ③ 被运输到细胞表面。(b) MHC 类 II 分子与内质网中的不变链 (Ii) 相关 ①。Ii-相关复合物被靶向到内吞泡腔 ②,在那里从内吞蛋白产生的肽类被装载到 MHC 类 II 分子上 ③,并转移到细胞表面 ④。